ОРИГИНАЛЬНОЕ ИССЛЕДОВАНИЕ
Своеобразие поморов Онежского полуострова и Зимнего берега в генетическом контексте севера Европы
1 Медико-генетический научный центр, Москва, Россия
2 Биобанк Северной Евразии, Москва, Россия
Своеобразие генофонда Русского Севера, давно став научным фактом, так и не получило содержательного объяснения. Резкие генетические отличия северных русских популяций не только от южных, но и друг от друга, нельзя объяснить лишь дрейфом генов. Расширение спектра изученных генофондов Русского Севера и их соседей может дать ключ к разгадке этого феномена. Целью работы было субтотальное исследование генофонда поморов Архангельской области (Онежского берега, Летнего берега, западного фрагмента Зимнего берега; n = 130) по панели 60 SNP-маркеров Y-хромосомы методами многомерного шкалирования и картографирования генетических расстояний. Популяции поморов резко различаются по частотам 14 выявленных гаплогрупп: каждая из гаплогрупп I1, R1a, N3 составляет по четверти общего генофонда поморов, по 8% I2-P37.2 и R1b, остальные гаплогруппы редки. Поморы Онежского берега оказались генетически схожими с широким кругом финноязычных народов СевероВосточной Европы и тех русских популяций, у которых есть мощный дославянский субстрат. Поморы Летнего берега близки к генофондам Скандинавии. Поморы Зимнего берега схожи лишь с единичными популяциями финнов и шведов. Ни одна из популяций поморов не имеет генетического сходства с населением Новгородчины, с которым традиционно связывают происхождение поморов. Генетические расстояния между популяциями поморов настолько велики, что сопоставимы с общим размахом изменчивости между восточными славянами, балтами и финно-уграми региона. Причинами столь ярко выраженного своеобразия популяций поморов наряду с дрейфом генов предположительно можно назвать разный дославянский субстрат, лежащий в основе генофонда каждой популяции, а также более поздние потоки генов.
Ключевые слова: генофонд, геногеография, Y-хромосома, Y-SNP, Русский Север, поморы, Фенноскандия
Финансирование: исследование выполнено при поддержке грантов РФФИ №20-09-00479_а (экспедиционное обследование, статистический анализ, генотипирование), РНФ №21-14-00363 (экспедиционное обследование, пробоподготовка, написание статьи), Государственного задания Министерства науки и высшего образования РФ для Медико-генетического научного центра им. академика Н. П. Бочкова (картографический анализ, интерпретация результатов).
Благодарности: авторы благодарят всех участников экспедиционного обследования (доноров образцов), Администрацию и Министерство здравоохранения Архангельской области — за организационную поддержку и содействие в проведении экспедиционного обследования, АНО «Биобанк Северной Евразии» — за предоставление коллекций ДНК.
Вклад авторов: Е. В. Балановская — руководство, остальные авторы — участники экспедиционного обследования поморов; Г. Ю. Пономарев, В. Ю. Пылёв — генотипирование Y-SNP маркеров; Анастасия Т. Агджоян, В. С. Окованцев, Анна Т. Агджоян — статистический и картографический анализ; Е. В. Балановская, Анастасия Т. Агджоян и В. С. Окованцев — дизайн исследования и написание статьи.
Соблюдение этических стандартов: исследование одобрено этическим комитетом Медико-генетического научного центра имени Н. П. Бочкова (протокол № 1 от 29 июня 2020 г.).
Для корреспонденции: Владимир Сергеевич Окованцев
ул. Москоречье, д. 1, 115522, г. Москва, Россия; ur.xednay@vokoavov
Изучение генетической истории русского народа охватывает все более широкий круг как русских популяций, так и их соседей [1–18]. На возникающей панораме ярко выделяется «белое пятно» — самые северные русские популяции Беломорья, поморы. Их изучение важно не только в качестве модели эволюции популяций на самой периферии этнического ареала. Дело в том, что Беломорская периферия крайне своеобразна: вся жизнь поморов, связанная с морем, формировала не только особый быт, но и особые пути взаимодействия с населением Северной и Северо-Восточной Европы.
Беломорскими поморами называли русских побережья Белого моря, которые промышляли охотой на морского зверя, рыболовством в открытом море, морской торговлей, отличались многими особенностями быта и сохранением черт древнерусской культуры [19]. С новгородской (верховой) колонизацией связаны первые упоминания о постоянных русских поселениях на беломорском побережье в конце XIII — начале XIV в. [20–22]. Согласно летописям, переселенцы встретили на этих землях финноязычное население — племена чуди заволочской, часто ассоциирующейся с вепсами. Но заселение региона началось в мезолите, т. е. около 8 тыс. лет назад. В неолите фиксируют следы двух близких археологических культур — каргопольской и беломорской. Новую волну пришлого населения в эпоху бронзы (4–3 тыс. лет назад) связывают с финно-угорским населением (прежде всего, с саамским [23–28]), а последнюю славянскую волну мигрантов — с Новгородской колонизацией северных пределов.
Жизнь на северной периферии ареала русского народа (где в топонимике прочно сохранились следы дославянского населения), своеобразие культуры и особый тип хозяйства поморов позволяют предполагать, что они имели и своеобразный генофонд. Лишь небольшая выборка поморов (n = 28) была изучена ранее по крайне узким панелям маркеров Y-хромосомы и мтДНК [1, 2]. Исследователи рассматривали поморов в составе популяции «северных русских» в широком контексте населения Северной Евразии [1]. Особенности их генофонда объяснены возможной ассимиляцией урало- или балтоязычного населения славянским [2]. На сходство с финноязычным населением (причем больше с финно-пермским, чем с финно-волжским) указывают данные об аутосомном геноме поморов Мезенского района Архангельской области (n = 96) [3].
Более широкий круг популяций, а именно генофонд Русского Севера, охарактеризован по четырем системам маркеров (Y-хромосомы, мтДНК, аутосомных ДНКмаркеров и частот фамилий). В анализ включены популяции Пинежского, Лешуконского, Красноборского и Ленского районов Архангельской области [4], масштаб генетического своеобразия которых показан ранее [5]. По мтДНК и аутосомным ДНК-маркерам эти популяции сближаются с населением Северной Европы. Разнообразие «отцовских» линий выявляет сходство генофонда Русского Севера с населением обширной территории от Прибалтики до Печоры и обусловлено наследием древнейшего палеоевропейского населения. Геномные данные об этих популяциях были привлечены для анализа генетической истории балто-славянских народов. На картах генетических расстояний популяции Русского Севера формируют один из основных паттернов генофонда Европы [7]. Поиск генетических следов новгородской колонизации в генофонде Русского Севера, проведенный по широкогеномной панели аутосомного генома [8], показал отсутствие «новгородской» предковой компоненты на севере Архангельской области, а на ее юге (Красноборский и Ленский районы) вклад «новгородской» компоненты составил не более трети генофонда.
Даже столь краткий обзор изученности генофонда Русского Севера выявляет серьезную нехватку данных о его самой северной периферии: описаны либо «материковые» популяции Архангельской области, либо маленькая выборка поморов по столь узкой панели, которая несопоставима с современным спектром ДНК-маркеров.
Новые данные о генофонде популяций поморов, изученном по обширной панели маркеров, могут дать ключ к пониманию огромного генетического разнообразия и своеобразия популяций Русского Севера [4, 5, 7]. С учетом традиции патрилокальности у поморов [23] и высокой эффективности изучения «отцовских линий» [1, 9–11], целью данной работы было рассмотреть полиморфизм маркеров Y-хромосомы для решения двух задач: создания «генетических портретов» трех впервые изученных популяций поморов и поиска в их генофонде генетических следов новгородской колонизации.
МАТЕРИАЛЫ И МЕТОДЫ
Материалы
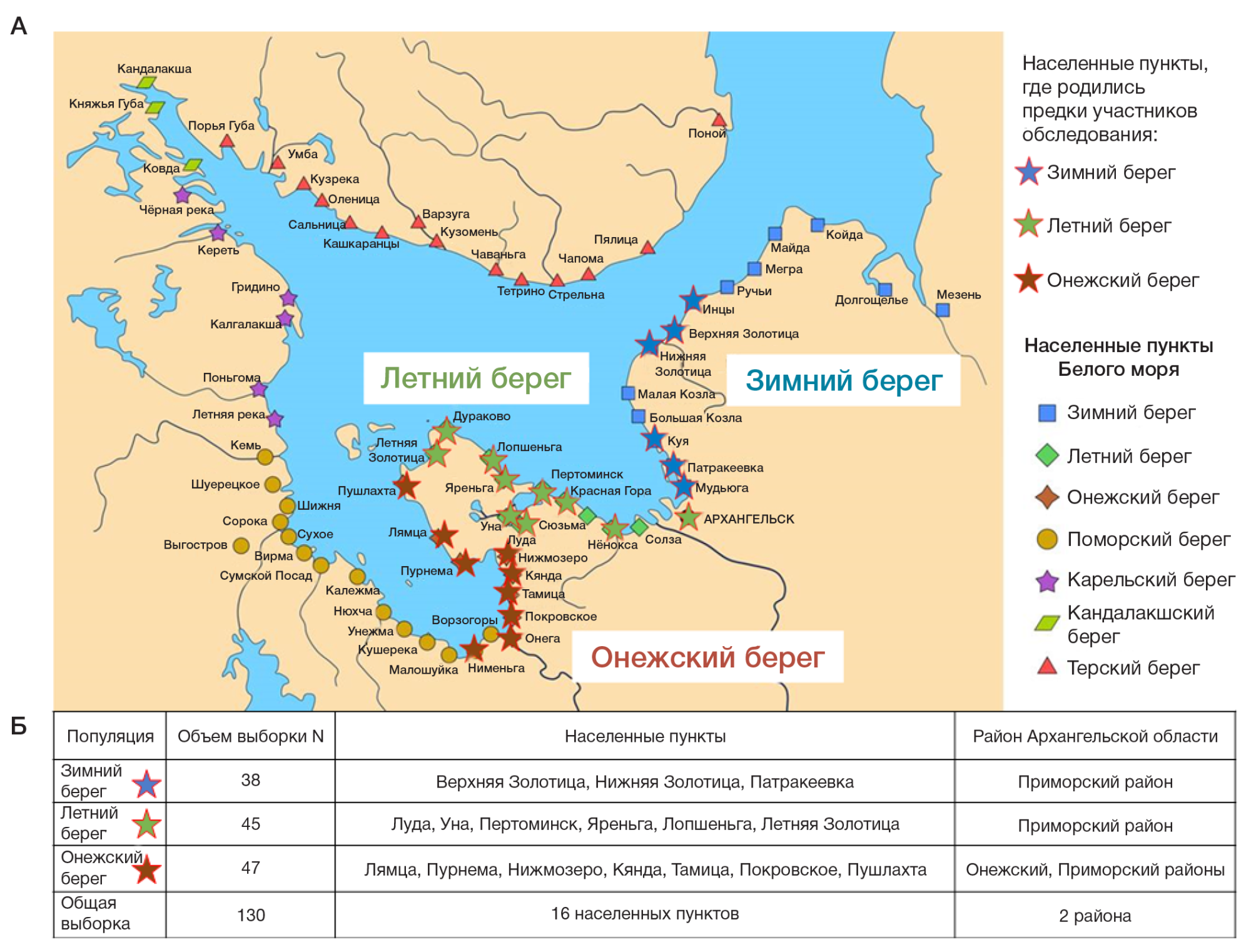
Коренное население побережья Белого моря детально изучено по современной панели маркеров Y-хромосомы (рис. 1). В ходе экспедиции 2021 г. субтотально обследованы населенные пункты Онежского полуострова (Онежский берег и Летний берег) и западного (Онежского) фрагмента Зимнего берега (рис. 1) (далее для краткости все популяции условно именуются «онежскими поморами»). Обследование проводили субтотально: образцы крови брали в селениях с компактным проживанием поморов практически у всех мужчин, соответствующих критериям включения в исследование: в выборку включали только тех неродственных между собой индивидов, чьи предки до третьего поколения принадлежали к данной популяции и считали себя русскими (или поморами). Для сравнения с популяциями коренного населения севера Европы привлечены литературная информация и неопубликованные данные АНО «Биобанк Северной Евразии».
Генотипирование
Тотальную ДНК выделяли из образцов венозной крови методом магнитных частиц на автоматизированной установке QIAsymphony (QIAGEN; Нидерланды). Генотипирование проводили по панели 60 Y-SNP маркеров методом ПЦР в реальном времени с использованием TaqMan-зондов и технологии OpenArray на амплификаторе QuantStudio 12 Flex (Thermo Fisher Scientific; США): D-M174, E-M35, E-M78, C-M217, C-F3791, C-F5481, C-F3918, C-M48, C-SK1066, C-M407, G-M201, G1-M285, G2-P15, G2FGC595, G2-M406, G2-P303, H-M69, I-M170, I-M253, I-P37.2, I-M223, J1-M267, J1-P58, J2-M172, J2-M12, J2-M67, J2-M9, L-M20, L-M317, T-M70, N-M231, N-M128, N-Y3205, N-M178, N-B211, N-M2118, N-CST10760, N-Z1936, N-F4205, N-B202, N-B479, O-P186, O-M119, O-P31, O-M122, O-P201, O-M134, Q-M242, R1a-M198, R1a-PF6202, R1a-Y2395, R1a-CTS1211, R1a-Z92, R1a-Z93, R1b-M343, R1b-Y13887, R1b-M269, R1b-L51, R1b-Z2105, R2-M124.
Статистический и картографический анализ
По данным о частотах 14 гаплогрупп Y-хромосомы, выявленных в трех популяциях поморов, рассчитывали матрицу попарных генетических расстояний М. Нея (в оригинальной программе DJ [5]), на основе которых построен график многомерного шкалирования в пакете Statistica 7.0. (StatSoft; США). Картографический анализ проводили с помощью оригинального программного пакета GeneGeo [29] по расширенному спектру 26 гаплогрупп Y-хромосомы, характерных для региона. Карты распространения 26 гаплогрупп строили по частотам из базы данных «Y-base» (разработанной под руководством О. П. Балановского) методом средневзвешенной интерполяции с радиусом влияния 800 км и третьей степенью весовой функции [29]. Алгоритм построения каждой карты генетических расстояний включал два этапа. На первом этапе для каждой из 26 гаплогрупп строили карту генетических расстояний от данной популяции поморов до интерполированных значений в каждой точке карты. На втором этапе по совокупности этих 26 карт рассчитывали средние генетические расстояния от данной популяции поморов до каждой точки карты. В результате для каждой популяции онежских поморов была создана карта, где показана степень генетического сходства изученной популяции поморов с каждой из популяций сравнения.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
Спектр гаплогрупп Y-хромосомы
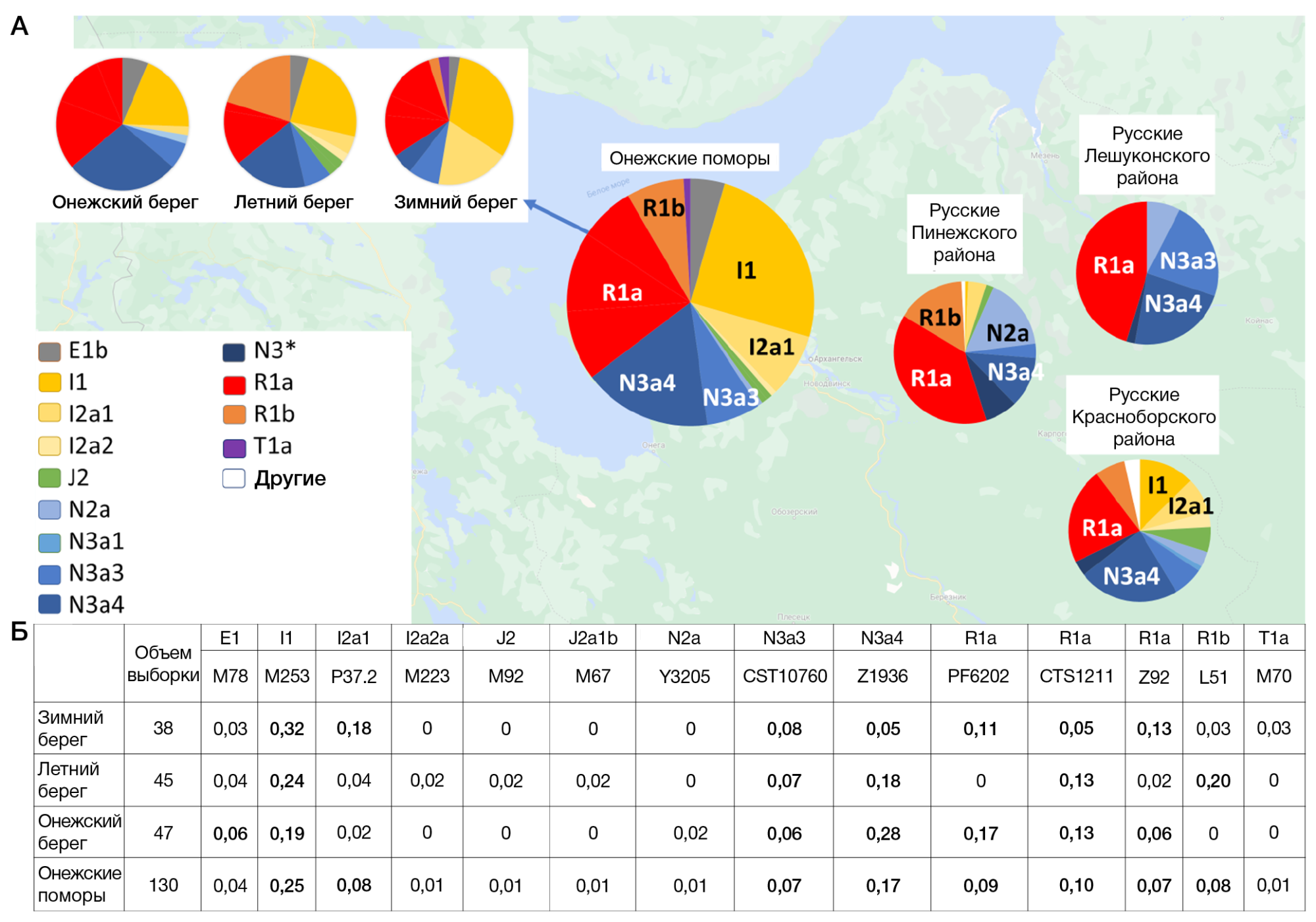
В генофондах трех популяций онежских поморов обнаружено 14 гаплогрупп Y-хромосомы (рис. 2): E-M78, I1-M253, I2-P37.2, I2-M223, J2-M92, J2-M67, N2a-Y3205, N3a3-CST10760, N3a4-Z1936, R1a-PF6202, R1a-CTS1211, R1a-Z92, R1b-L51, T1a-M70 (далее указаны краткие названия этих гаплогрупп). Самыми частыми оказались гаплогруппы I1, N3 и R1a, каждая из которых составляет около четверти общего генофонда онежских поморов (рис. 2). Гаплогруппа R1a представлена тремя ветвями (PF6202, CTS1211, Z92), гаплогруппа N3 — двумя (CST10760, Z1936). Следующими по убыванию частоты следуют гаплогруппы I2-P37.2 и R1b (по 8% генофонда в целом), остальные редки.
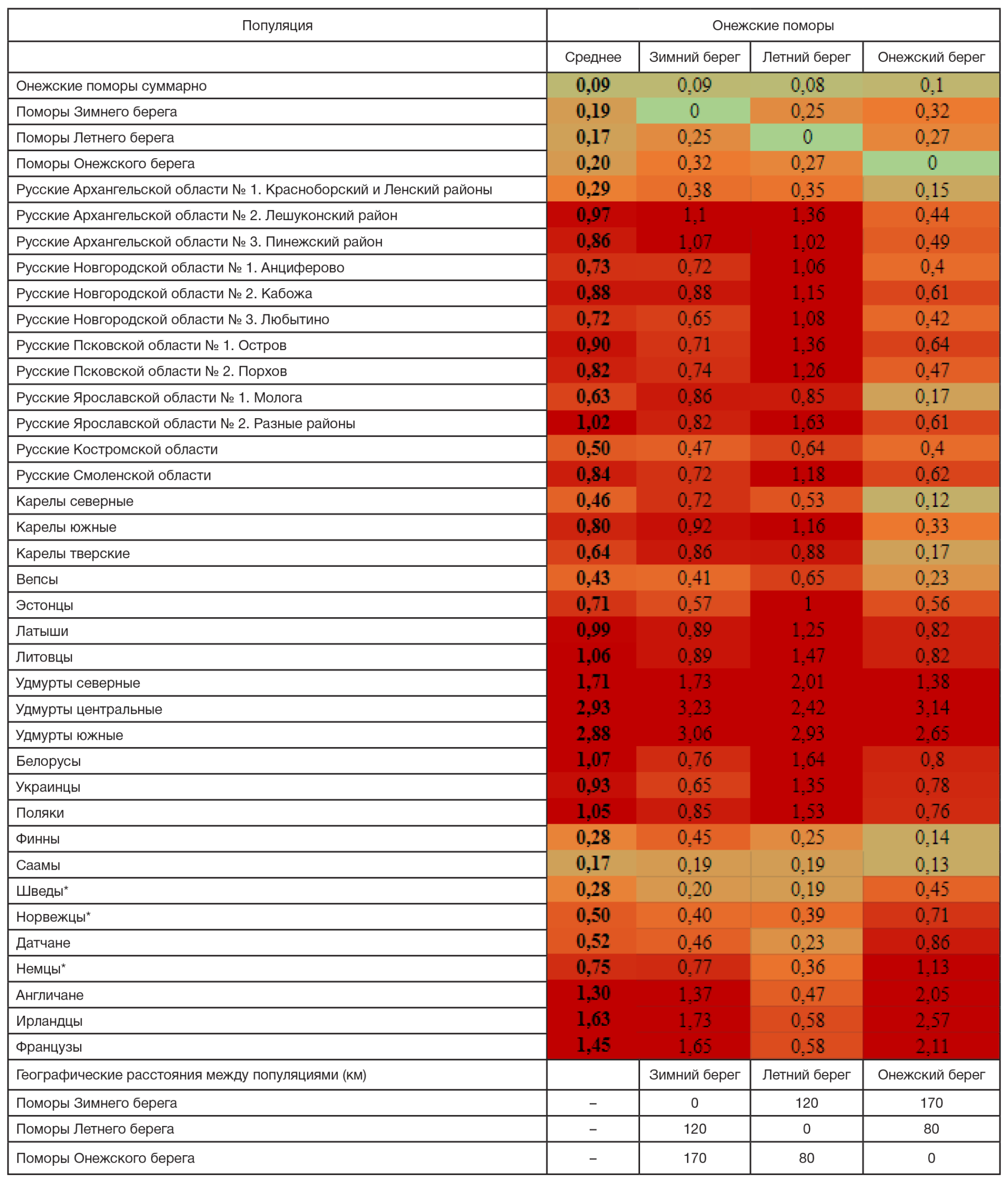
Вопреки географической близости трех популяций поморов (80–170 км; таблица), их генетические портреты заметно различаются, причем по четырем гаплогруппам (I2, N3a4, R1a, R1b) эти различия достоверны. Хотя у каждой из трех популяций поморов присутствуют не менее девяти гаплогрупп их «общего портрета», спектр гаплогрупп каждой популяции очень своеобразен. У поморов Зимнего берега снижена частота гаплогруппы N3a4 и повышена частота I2; в популяции Летнего берега не обнаружена гаплогруппа R1a-PF6202, характерная других популяций, но повышена частота R1b; в популяции Онежского берега высока частота N3a4, но низка I2 (рис. 2).
Прослеживаются убывание доли гаплогрупп I1 и I2a c востока (Зимний берег) на запад (Онежский берег) и противоположный тренд у гаплогрупп N3a3 и N3a4. Но такой «долготный» тренд отсутствует у гаплогрупп R1a (частота повышена в популяциях Зимнего и Онежского берегов) и R1b (максимум на Летнем берегу). Частота гаплогруппы R1a не только высока в генофондах Зимнего (29%) и Онежского берегов (36%), но и все три ветви R1a здесь встречены с частотой ≥ 5%. Однако на Летнем берегу частота R1a в два раза ниже и обнаружены только ветви R1a-CTS1211 (13%) и R1a-Z92 (2%). Снижение частоты гаплогруппы R1a и резкое возрастание частоты R1b (20%) только в популяции Летнего берега могут быть результатом и дрейфа генов, и их миграционного потока. Высока частота R1b и у русских Пинежского района Архангельской области (рис. 2), но там распространена иная ветвь R1b. У поморов обнаружена ветвь L51, более характерная для народов Северо-Западной, чем СевероВосточной Европы, и для установления ее источника (миграция или сохранение древнейшего генетического ландшафта региона) необходимо использование филогенетических подходов.
С другими русскими популяциями Архангельской области онежские поморы наиболее схожи по заметной роли гаплогрупп R1a и N3a4 (хотя в популяции Зимнего берега N3a4 редка). Резкое отличие «прибрежных» поморов от «материковых» популяций Архангельской области проявляется по частоте гаплогруппы I1: у онежских поморов она составляет в среднем четверть генофонда (25%), но для других северных русских не характерна (в Красноборской популяции — 12%, в Пинежской — 1%, в Лешуконской не обнаружена).
Положение онежских поморов в генетическом пространстве Северной Европы
При анализе степени генетического сходства (таблица) прежде всего удивляет, сколь велики генетические расстояния между популяциями поморов (d = 0,28) вопреки их географической близости и культурной общности. Более того, расстояние от онежских поморов до других русских популяций почти в три раза больше (d = 0,76), но есть и очень выразительные исключения (таблица). Наиболее близкой к генофонду онежских поморов (d = 0,29) оказалась географически удаленная популяция (около 500 км) русских Красноборского и Ленского районов Архангельской области. При этом она чрезвычайно близка лишь к поморам Онежского берега (d = 0,15), но генетически далека от Летнего (d = 0,33) и Зимнего (d = 0,38) берегов.
Из других русских популяций относительное генетическое сходство с поморами обнаруживают лишь русские Костромской области (d = 0,50) и популяция Мологи Ярославской области (d = 0,63). При этом к генофонду Мологи близки только поморы Онежского берега (d = 0,17), в то время как генофонды Летнего и Зимнего от нее крайне далеки (d = 0,85). Ранее было показано, что среди ярославских популяций именно Молога сохранила явный генетический след дославянского населения (предположительно мери) [9].
Среди иных народов России к онежским поморам наиболее генетически близки вепсы (d = 0,43) и северные карелы (d = 0,46), причем вновь лишь за счет поморов Онежского берега (d = 0,23 и d = 0,12 соответственно). Оказывается, что поморы Онежского берега генетически ближе к финноязычным соседям, чем к другим популяциям поморов (d = 0,28). А вот представители восточного крыла финноязычных народов — удмурты — оказались самыми генетически далекими от поморов (d = 2,50), что противоречит выводу [3] о сходстве генофонда поморов и финно-пермских народов.
Однако наибольшее генетическое сходство онежские поморы обнаруживают с народами зарубежной Северной Европы (таблица): генетическое расстояние от поморов до шведов и финнов (d = 0,28) такое же, как среднее расстояние между популяциями поморов (d = 0,28), а расстояние до генофонда саамов в два раза меньше (d = 0,14). Причем прослеживается любопытный тренд: расстояние до генофондов финнов и саамов уменьшается в четыре раза по мере движения на запад — от Зимнего берега к Онежскому. Но генетические расстояния до скандинавов (датчан, норвежцев, шведов) следуют противоположному тренду: расстояние до Онежского берега в два раза больше, чем до Летнего и Зимнего, которые одинаково близки к скандинавам. При этом, если поморы Зимнего берега близки лишь к скандинавам из всего пула рассмотренных популяций, то поморы Летнего берега обнаруживают определенное генетическое сходство также и с широким кругом европейских популяций — от немцев (d = 0,36) до ирландцев (d = 0,65).
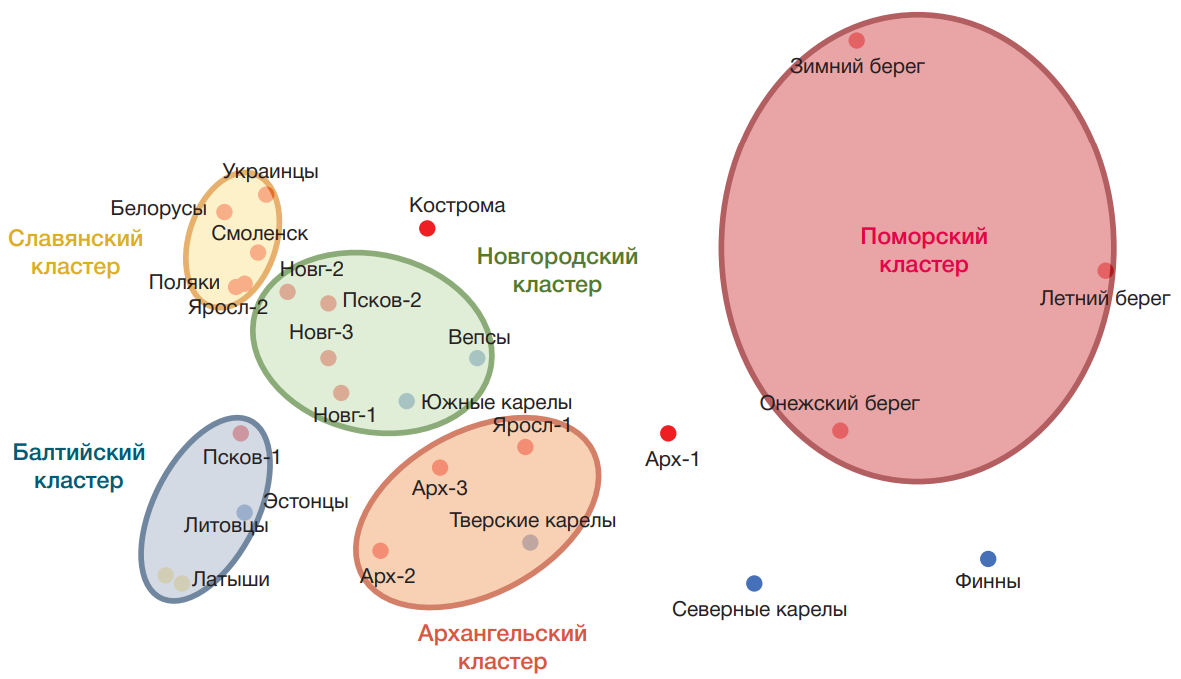
В генетическом пространстве многомерного шкалирования (рис. 3) (график построен по 14 «поморским» гаплогруппам Y-хромосомы) выделяются пять кластеров (им даны весьма условные названия); для каждого рассчитаны средние расстояния между популяциями (d͞). «Славянский» кластер (d͞ = 0,05) включил белорусов, украинцев, поляков и русских Смоленской и Ярославской областей. К нему близок «Новгородский» кластер (d͞ = 0,06), объединивший все три популяции Новгородской области и Порхов Псковской области, который входил в Новгородские земли. К ним присоединились также финноязычные популяции — вепсы и южные карелы. – «Прибалтийский» кластер (d͞ = 0,04) включил всех прибалтов: латышей, литовцев и эстонцев, а также популяцию Псковской области (Остров). «Архангельский» кластер (d͞ = 0,09) объединил популяции Пинежского и Лешуконского районов Архангельской области с Мологой Ярославской области и тверскими карелами.
Поморы образовали свой собственный обширный кластер — расстояния между популяциями поморов (d͞ = 0,28) почти в пять раз больше, чем среднее расстояние в пределах других кластеров (d͞ = 0,06), а площадь «Поморского» кластера лишь немногим меньше суммы всех четырех кластеров групп сравнения, включивших финноязычные, балтоязычные и славянские популяции. Но подчеркнем, что при столь огромных различиях между поморскими популяциями все они заняли свое собственное «поморское» место в генетическом пространстве Северо-Восточной Европы.
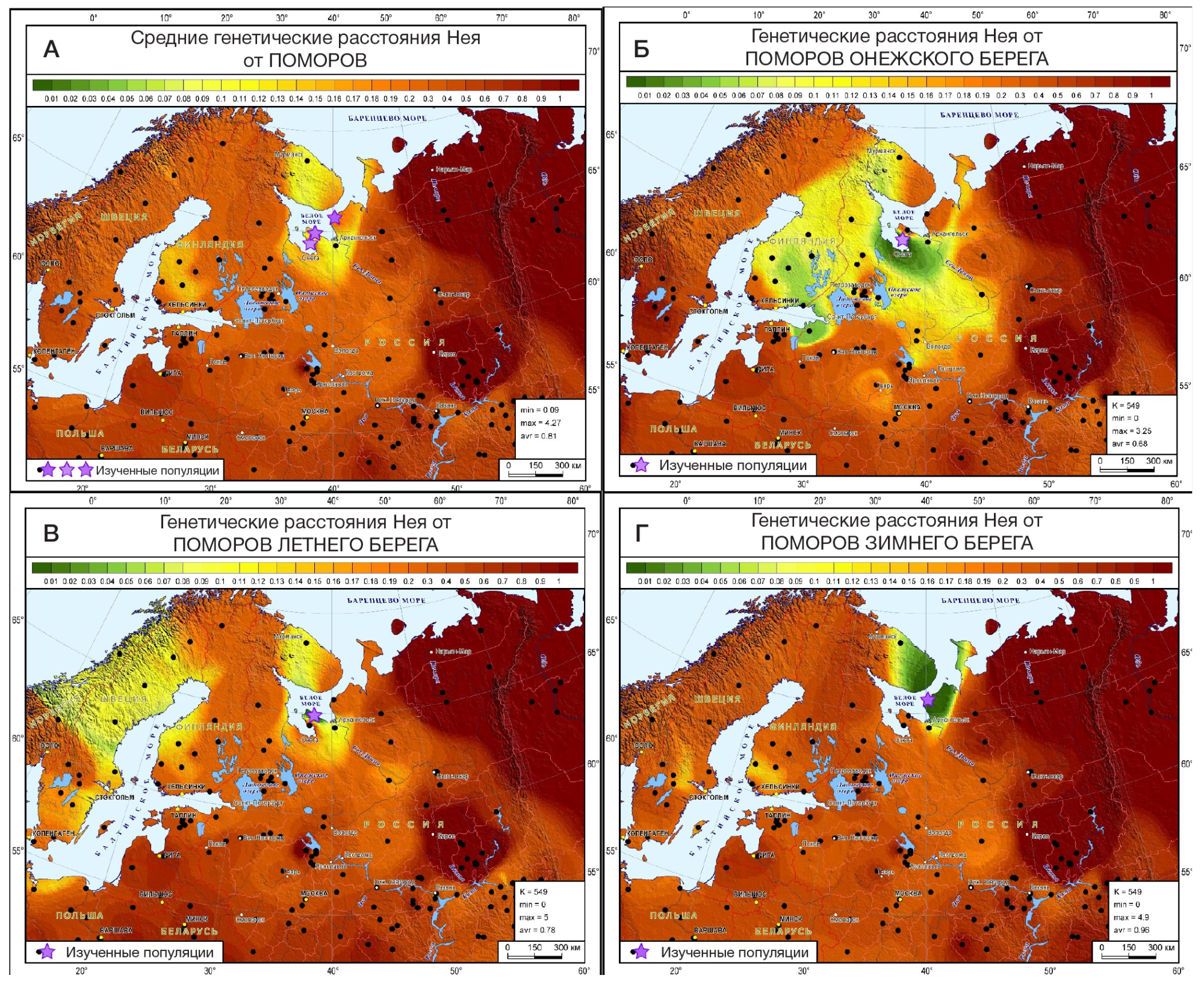
Более точно определить регионы, с которыми схожи генофонды поморов, и значительно расширить круг популяций сравнения помогают карты генетических расстояний (рис. 4), рассчитанные по 26 гаплогруппам Y-хромосомы, характерным для всего региона.
Суммарный генофонд поморов (рис. 4А) генетически близок к южной части Финляндии, а не более северной, представленной саамами.
Генофонд Онежского берега (рис. 4Б) выявляет обширную область генетического сходства: она охватывает почти всю Финляндию на западе, с востока четко очерчена Северной Двиной и Сухоной, на юге и юго-западе России доходит до Ярославской и Ленинградской областей. В эту область сходства входят и финноязычные народы (вепсы, ижора, ингерманландцы, карелы, финны), и те русские популяции, в генофонде которых прослеживается существенный вклад дославянского населения.
Генофонд Летнего берега (рис. 4В) обнаружил наибольшее сходство с далекими шведами и норвежцами, слабее оно выражено с саамами, представляющими самый север Скандинавии.
Генофонд Зимнего берега (рис. 4Г) относительно генетически близок лишь к нескольким популяциям финнов и шведов. Это единственная популяция поморов, для которой можно допустить, что ее генетический портрет в значительной степени сформирован дрейфом генов. Однако популяция Зимнего берега пока представлена лишь ее «прионежской» частью (рис. 1). Сейчас ведется изучение генофонда всего Зимнего берега, что позволит вскоре дать обоснованное заключение о его генетической истории.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Все три рассмотренные популяции поморов не только географически близки (рис. 1): их хозяйственнокультурный тип, в отличие от оседлых земледельцев, предполагает передвижения по морю на большие расстояния. Поэтому разумно было предполагать, что различия между их генофондами будут крайне невелики. При экспедиционном обследовании ставилась задача сформировать субтотальную выборку, чтобы уловить даже незначительные генетические различия между тремя поморскими берегами: были обследованы все поселения с компактным проживанием онежских поморов (рис. 1). И хотя анализируемые выборки невелики (37–48 человек), они надежны, так как репрезентативно представляют генеральную совокупность благодаря субтотальности обследования и отражают реальность, а не выборочную ошибку. Даже при столь небольших выборках различия по четырем гаплогруппам (I2, N3a4, R1a, R1b) из 14 выявленных оказались достоверны, хотя этот вид анализа основан на предположении, что выборки сделаны из бесконечной генеральной совокупности индивидов. Поэтому анализ достоверности различий неприменим к субтотальным исследованиям малых популяций: именно субтотальные выборки дают наиболее точный портрет популяции и не требуют дополнительной оценки достоверности различий.
Вопреки изложенным выше ожиданиям оказалось, что каждая из трех поморских популяций имеет ярко выраженный уникальный генетический портрет. Поморы Онежского берега генетически близки и к финноязычным народам России и Финляндии, и к русским популяциям Архангельской области. Более того, они генетически ближе к финноязычным соседям, чем к другим онежским поморам. У поморов Летнего берега обнаружено генетическое сходство только с населением Скандинавии. Наконец, у поморов Зимнего берега практически нет схожих генофондов, за исключением некоторой близости к финнам и шведам. Различия между тремя популяциями поморов столь велики, что лишь немногим уступают всей совокупности различий между рассмотренными популяциями (рис. 3) западных и восточных славян, балтов и финноязычных народов. И при этом все три популяции поморов занимают свое собственное «поморское» место в генетическом пространстве столь широкого круга популяций.
Связать столь уникальное своеобразие генофондов поморов с действием лишь дрейфа генов невозможно. Дрейф генов независимо действует на разные гаплогруппы. Поэтому «дрейфующая» популяция по одному из маркеров может стать схожей с одной из сравниваемых популяций, но по другой гаплогруппе она окажется близка к совсем другой популяции, по третьей гаплогруппе — к третьей популяции и т. д. Поэтому, когда мы проводим анализ генетических расстояний суммарно по всей совокупности генетических маркеров, такая «дрейфующая» популяция вне зависимости от ее реального происхождения оказывается непохожей ни на одну из групп сравнения.
Данная модель в какой-то мере может объяснять своеобразие генофонда поморов Зимнего берега. Но окончательный вывод о ней можно будет сделать лишь после анализа генетических портретов других популяций обширного Зимнего берега (рис. 1). Несомненно, что дрейф генов был важным фактором в генетической истории всех популяций поморов, которые в течение последних поколений становились все менее многочисленными. Однако дрейфу генов не удалось стереть генетическую память о том, что в основе их генофондов имелся различный субстрат. Для поморов Онежского берега он общий с широким кругом западного ствола финноязычного населения севера Европы, тогда как поморы Летнего берега схожи только с населением Скандинавии. Полногеномные исследования позволят верифицировать гипотезы их разного происхождения, давних миграционных потоков генов, сформировавших их генофонды. Однако результаты изучения полиморфизма Y-хромосомы (наиболее стабильной части генофонда вследствие патрилокальности поморов) прямо указывают, что причина генетического своеобразия популяций онежских поморов кроется в различном генетическом субстрате, в различном происхождении, хотя эти различия и были перекрыты мощным дрейфом генов.
Второй важный вопрос — о генетическом сходстве поморов и новгородцев. Среднее генетическое расстояние между этими популяциями (d = 0,77) оказалось таким же (d = 0,76), как и расстояние между генофондами поморов и остальных рассмотренных русских популяций (таблица). Генетические отличия новгородцев велики даже от поморов Онежского берега (d = 0,48), но от популяций других поморов они вообще огромны (от Зимнего берега d = 0,75, от Летнего берега d = 1,09). Ранее нами был сделан вывод об отличиях аутосомного генома новгородцев от русских севера Архангельской области [8]. Теперь мы видим выраженные различия между генофондами новгородцев и поморов и по маркерам Y-хромосомы. Оба результата противоречат мнению, что генофонд Русского Севера был сформирован новгородской экспансией. Однако это далеко не единственный случай в мировой культуре, когда внутренняя колонизация выражалась в расширении власти и экономического влияния, но не приводила к решительному изменению генофонда.
Эти и другие результаты изучения коренного населения Европы [4–5, 8–12, 30] убедительно показывают, что многие представления об его истории, сложившееся на базе данных гуманитарных наук, далеко не всегда могут быть интерполированы на генофонд.
ВЫВОДЫ
1. Изучен полиморфизм Y-хромосомы в трех популяциях поморов Белого моря: Онежского, Летнего и Зимнего берегов. Анализ субтотальных выборок неродственных индивидов из всех пунктов с компактным проживанием поморов позволил создать надежные генетические портреты трех популяций поморов. 2. Исследование генофонда поморов по широкой панели маркеров Y-хромосомы выявило 14 гаплогрупп, из которых по четырем (I2, N3a4, R1a, R1b) обнаружены достоверные различия, несмотря на небольшие объемы выборок (37–48 человек). Различия генетических портретов поморов складываются за счет своеобразия генофонда каждой из популяций: у поморов Зимнего берега снижена частота гаплогруппы N3a4 и повышена частота I2; в популяции Летнего берега не обнаружены ветви гаплогруппы R1a, характерные для двух других популяций, но повышена частота R1b; в популяции Онежского берега высока частота N3a4, но низка I2. Особенности спектра и частот гаплогрупп создают уникальность генетического портрета каждой популяции поморов. 3. Каждой из трех популяций поморов присущ собственный спектр генетически близких популяций. Поморы Онежского берега генетически схожи с широким кругом финноязычных народов севера Европы и с некоторыми русскими популяциями, в генофонде которых выражен вклад дославянского населения. Поморы Летнего берега обнаружили сходство только с населением Скандинавии, что можно объяснить общим палеоевропейским субстратом или более поздними взаимодействиями скандинавов и поморов. Для поморов Зимнего берега зафиксирована некоторая генетическая близость лишь к двум популяциям Финляндии и Швеции. 4. Генетические расстояния между популяциями поморов оказались сопоставимы с общим размахом изменчивости между восточными славянами, балтами и финно-уграми региона. Причиной этого может быть (в совокупности с дрейфом генов) разный субстрат, лежащий в основе генофонда каждой популяции, поскольку трудно предположить столь разные миграционные связи у столь географически, этнически и культурно близких популяций. 5. Ни одна из трех популяций онежских поморов не проявляет заметного генетического сходства с коренным населением Новгородчины, что указывает на отсутствие генетических следов демической экспансии при новгородской колонизации Русского Севера.
- Wells RS, Yuldasheva N, Ruzibakiev R, Underhill PA, Evseeva I, BlueSmith J et al. The Eurasian Heartland: A continental perspective on Y-chromosome diversity. Proc Natl Acad Sci USA. 2001; 98 (18): 10244–9.
- Malyarchuk BA. Differentiation of the Mitochondrial Subhaplogroup U4 in the Populations of Eastern Europe, Ural, and Western Siberia: Implication to the Genetic History of the Uralic Populations. Russian Journal of Genetics. 2004; 40 (11): 1281–7.
- Khrunin AV, Khokhrin DV, Filippova IN, Esko T, Nelis M, Bebyakova NA, et al. A Genome-Wide Analysis of Populations from European Russia Reveals a New Pole of Genetic Diversity in Northern Europe. PLoS One. 2013; 8 (3): e58552.
- Балановская Е. В., Пежемский Д. В., Романов А. Г., Баранова Е. Е., Ромашкина М. В., Агджоян А. Т. и др. Генофонд Русского Севера: славяне? Финны? Палеоевропейцы? Вестник Московского университета. 2011; (3): 27–58.
- Balanovsky O, Rootsi S, Pshenichnov A, Kivisild T, Churnosov M, Evseeva I et al. Two Sources of the Russian Patrilineal Heritage in Their Eurasian Context. The American Journal of Human Genetics. 2008; 82 (1): 236–50.
- Triska P, Chekanov N, Stepanov V, Khusnutdinova EK, Kumar GPA, Akhmetova V, et al. Between Lake Baikal and the Baltic Sea: genomic history of the gateway to Europe. BMC Genetics. 2017; 18 (1): 110.
- Балановский О. П. Генофонд Европы. М.: Товарищество научных изданий КМК, 2015; 354 с. [цитируется по 21 май 2022 г.]. Доступно по ссылке: http://генофонд.рф/wp-content/ uploads/Balanovskiy_2015-.pdf.
- Балановская Е. В., Черневский Д. К., Балановский О. П. Своеобразие Новгородского генофонда в контексте народонаселения европейской части России. Вестник НовГУ. 2021; 124 (3): 51–7.
- Chukhryaeva M, Pavlova E, Napolskich V, Garin E, Klopov A, Temnyatkin S, et al. Is there a Finno-Ugric component in the gene pool of Russians from Yaroslavl oblast? Evidence from Y-chromosome. Russian Journal of Genetics. 2017; 53: 388–99.
- Fechner A, Quinque D, Rychkov S, Morozowa I, Naumova O, Schneider Y, et al. Boundaries and clines in the West Eurasian Y-chromosome landscape: insights from the European part of Russia. Am J Phys Anthropol. 2008; 137 (1): 41–7.
- Malyarchuk B, Derenko M, Grzybowski T, Lunkina A, Czarny J, Rychkov S, et al. Differentiation of mitochondrial DNA and Y chromosomes in Russian populations. Hum Biol. 2004; 76 (6): 877–900.
- Балановская Е. В., Балановский О. П. Русский генофонд на Русской равнине. М.: Луч, 2007; 412 с.
- Булыгина Е. А., Соловьев В. Д. Генетическое и лингвистическое разнообразие четырнадцати популяций Евразии. Ученые записки Казанского университета. 2012; 154 (4): 175–87.
- Губина М. А., Бабенко В. Н., Дамба Л. Д., Пономарева М. Н., Коновалова Н. А., Воевода М. И. Полиморфизм митохондриальной ДНК у староверов Сибири. Генетика. 2014; 50 (6): 729–44. [цитируется по 6 сентябрь 2022 г.]. Доступно по ссылке: http://naukarus.com/polimorfizm-mitohondrialnoydnk-u-staroverov-sibiri.
- Rubinstein S, Dulik MC, Gokcumen O, Zhadanov S, Osipova L, Cocca M, et al. Russian Old Believers: genetic consequences of their persecution and exile, as shown by mitochondrial DNA evidence. Hum Biol. 2008; 80 (3): 203–37.
- Belyaeva O, Bermisheva M, Khrunin A, Slominsky P, Bebyakova N, Khusnutdinova E, et al. Mitochondrial DNA variations in Russian and Belorussian populations. Hum Biol. 2003; 75 (5): 647–60.
- Zhernakova DV, Brukhin V, Malov S, Oleksyk TK, Koepfli KP, Zhuk A, et al. Genome-wide sequence analyses of ethnic populations across Russia. Genomics. 2020; 112 (1): 442–58.
- Лимборская С. А., Вербенко Д. А., Хрунин А. В., Сломинский П. А., Бебякова Н. А. Этническая геномика: анализ геномного полиморфизма популяций Архангельской области. Вестник Московского университета Серия 23 Антропология. 2011; (3): 100–19.
- Власова И. В., Александров В. А., Полищук Н. С., редакторы. Русские. М.: Наука, 1999; 827 с.
- Макаров Н. А. Колонизация северных окраин Древней Руси в XI–XIII вв: По материалам археол. памятников на волоках Белозерья и Поонежья. М.: Науч.-изд. центр «Скрипторий», 1997; 368 с.
- Макаров Н. А. Археологическое изучение Северо-Восточной Руси: колонизация и культурные традиции. Вестник Российской Академии Наук. 2009; 79 (12): 1068–79.
- Makarov N. Rural Settlement and Landscape Transformations in Northern Russia, A.D. 900–1300. In: Hines J, Lane A, Redknap M, editors. Land, Sea and Home: Settlement in the Viking Period. London: Routledge, 2004; 55–74 p.
- Бернштам Т. А. Народная культура Поморья в XIX–начале XX в: этнографические очерки. Ленинград: Наука. Ленинградское отделение, 1983; 233 с.
- Бернштам Т. А. Поморы: Формирование группы и система хоз-ва. Ленинград: Наука Ленинградское отделение, 1978; 176 с.
- Перевозчиков И. В., Маурер А. М., Бацевич В. А., Локк К. Э. Обобщенный фотопортрет поморов (в связи с трехсотлетним юбилеем со дня рождения М. В. Ломоносова). Вестник Московского университета. Серия 23 Антропология. 2011; (3): 59–67.
- Пермиловская А. Б. Русский Север — хранитель «генофонда» русской этнокультурной традиции. В книге: Фурсова Е. Ф., Пермиловская А. Б., Черных А. В., Зуев А. С., Люцидарская А. А., Майничева А. Ю., и др. Сибирь и Русский Север. Новосибирск: Институт археологии и этнографии Сибирского отделения Российской академии наук, 2014; с. 7–42.
- Макаров Н. А. Русский Север: таинственное средневековье. М., 1993; 190 с.
- Пыжова А. Н. Этническая идентичность поморов Архангельской области через призму основных антропологических подходов. Арктика и Север. 2012; (7): 37–44.
- Кошель С. М. Геоинформационные технологии в геногеографии. В книге: И. К. Лурье, В. И. Кравцова, редакторы. Современная географическая картография. М.: Дата+, 2012; с. 158–66.
- Völgyi A, Zalán A, Szvetnik E, Pamjav H. Hungarian population data for 11 Y-STR and 49 Y-SNP markers. Forensic Sci Int Genet. 2009; 3 (2): e27–28.