ОРИГИНАЛЬНОЕ ИССЛЕДОВАНИЕ
Генотипическая характеристика штаммов Bordetella pertussis — кандидатов для получения коклюшного компонента вакцинных препаратов (сообщение I)
1 Московский научно-исследовательский институт эпидемиологии и микробиологии имени Г. Н. Габричевского, Москва, Россия
2 Научный центр экспертизы средств медицинского применения Минздрава России, Москва, Россия
3 Российский национальный исследовательский медицинский университет имени Н. И. Пирогова, Москва, Россия
Вакцинация является эффективным средством предупреждения заболевания коклюшной инфекцией. Целью работы было усовершенствование вакцинных препаратов в РФ, в том числе актуализация производственных вакцинных штаммов. Изучали штаммы B. pertussis, выделенные в г. Москве, Воронежской, Новосибирской, Ульяновской и Челябинской областях, и восемь производственных штаммов для адсорбированной коклюшно-дифтерийно-столбнячной (АКДС) вакцины. Для генотипирования использовали мультилокусное антигенное сиквенс-типирование (MAST) и полногеномное мультилокусное сиквенс-типирование (wgMLST). Изучены культурально-морфологические, ферментантивные, серологические и генотипические свойства штаммов B. pertussis — кандидатов в производственные вакцинные штаммы и проведен сравнительный анализ генотипических свойств этих штаммов и производственных вакцинных штаммов B. pertussis для вакцины АКДС. Кандидатные штаммы являются представителями четырех генотипов — ptxА1/ptxВ2/ptxС2/ptxР3/fim2-2/fim3-2/prn2, ptxА1/ptxВ2/ptxС2/ptxР3/fim2-2/fim3-2/prn9, ptxА1/ptxВ2/ptxС2/ptxР3/fim2-1/fim3-1/prn1 и ptxА1/ptxВ2/ptxС2/ptxР3/fim2-2/fim3-1/prn2. Производственные вакцинные штаммы принадлежали к другим шести генотипам: ptxА2/ptxВ1/ptxС1/ptxР1/ fim2-1/fim3-1/prn1, ptxА2/ptxВ2/ptxС1/ptxР2/fim2-1/fim3-1/prn1, ptxА4/ptxВ1/ptxС1/ptxР2/fim2-1/fim3-1/prn1, ptxА2/ptxВ2/ptxС1/ptxР1/fim2-1/fim3-1/prn1, ptxА4/ptxВ2/ptxС1/ptxР2/fim2-1/fim3-1/prn1 и ptxА1/ptxВ2/ptxС1/ptxР1/fim2-1/fim3-1/prn1. С помощью wgMLST установлена принадлежность всех кандидатных штаммов B. pertussis к ST2.
Ключевые слова: генотипирование, Bordetella pertussis, вакцинные препараты, мультилокусное антигенное сиквенс-типирование
Вклад авторов: О. Ю. Борисова — молекулярно-генетические исследования, анализ данных, анализ литературы, подготовка рукописи; И. Ю. Андриевская — молекулярно-генетические исследования, анализ данных, подготовка рукописи; А. С. Пименова, Н. Т. Гадуа, И. А. Чагина, И. А. Алексеева — микробиологические исследования, подготовка рукописи; А. Б. Борисова, Л. И. Кафарская — анализ литературы, анализ данных, подготовка рукописи; А. В. Чаплин — биоинформатический и филогенетический анализ, подготовка рукописи.
Для корреспонденции: Ольга Юрьевна Борисова
ул. Адмирала Макарова, д. 10, 125212, г. Москва, Россия; ur.liam@avosirobglo
Коклюшная инфекция является опасным, иногда смертельным заболеванием — в первую очередь, для новорожденных и детей первых месяцев жизни [1–3]. Актуальность исследования коклюша обусловлена ростом случаев заболеваемости, протеканием инфекции в тяжелых клинических формах, особенно у детей до одного года, а также наличием летальных исходов. В 2023 г. в РФ зарегистрировано 52 727 случаев коклюша (показатель заболеваемости — 36,2 на 100 тыс. населения), что в 16,4 раза выше уровня заболеваемости в 2022 г. (2,2 на 100 тыс. населения), и в 7,5 раз выше среднемноголетнего показателя заболеваемости (4,8 на 100 тыс. населения).
Из них более 80% были дети до 14 лет (данные Референсцентра по мониторингу за коклюшем и дифтерией в ФБУН МНИИЭМ им. Г. Н. Габричевского Роспотребнадзора по анализу формы № 2 «Сведения об инфекционных и паразитарных заболеваниях») [4–6].
Плановая вакцинация против коклюша, проводимая на первом году жизни ребенка, приводит к выработке у него поствакцинального иммунитета и невосприимчивости к инфекции [1, 3]. К 6–7 годам поствакцинальный иммунитет ослабевает. Поступление в школу связано с новыми контактами в детских коллективах, в которых возможно присутствие инфицированных детей. В таких коллективах наиболее высока вероятность заболевания непривитых, а также детей, утративших поствакцинальный иммунитет (последние, как правило, переносят коклюш в легкой форме с нетипичным кашлем) [7–9]. Показано, что 7–17% случаев продолжительного кашля у подростков связаны с инфекцией B. pertussis. По данным разных авторов, именно дети школьного возраста в 75% случаев становятся источниками инфекции B. pertussis для детей первого года жизни [9, 10]. Поэтому вакцинация является эффективным средством предупреждения заболевания коклюшной инфекцией у детей и особенно младенцев.
Рост заболеваемости коклюшем вызывает озабоченность специалистов относительно эффективности существующей стратегии профилактики [11, 12]. Наиболее вероятны следующие причины роста заболеваемости коклюшем: большое число случаев отказа родителей от вакцинации своих детей в возрасте до года и, следовательно, запаздывание начала прививок [2, 6]; накопление неиммунных лиц среди детей старших возрастных групп; утрата специфического иммунитета у взрослых [10, 13]; генотипическая изменчивость возбудителя под селективным давлением вакцинации [14–23]; распространение B. pertussis бессимптомными носителями [24]. Кроме того, внедрение в широкую практику ПЦР-исследований позволило значительно увеличить число выявленных случаев коклюша, в том числе у лиц с легкими, стертыми формами инфекции, а также при обследовании групповых случаев заболевания в очагах (неопубликованные данные Референс-центра по мониторингу за коклюшем и дифтерией.
Уже более 60 лет для вакцинации используют цельноклеточный коклюшный компонент комбинированной вакцины АКДС. Однако при своей высокой эффективности цельноклеточная вакцина обладает реактогенностью, что в свою очередь предопределило разработку бесклеточных вакцин, введенных в мировую практику здравоохранения со второй половины 1990-х гг. Такие вакцины содержат от 1 до 5 очищенных коклюшных антигенов (коклюшный токсин (PT), филаментозный геммаглютинин (FHA), пертактин (PRN), фимбрии 2-го и 3-го типа (Fim2 и Fim3)). Тем не менее, вакцинация населения не предотвращает в полной мере распространение коклюша, которое отмечают сейчас во многих странах мира. С 2000-х гг. на фоне широкого применения бесклеточной коклюшной вакцины в странах Европы, Японии, США, Австралии заболеваемость коклюшем стала постепенно расти, отмечались эпидемические вспышки [https://www.who.int/health-topics/pertussis#tab=tab_1]. Как считает ряд авторов, существует корреляция между применением бесклеточных вакцин и ростом заболеваемости коклюшем, что поднимает целый ряд вопросов относительно эффективности данных вакцин и их способности контролировать заболеваемость [11, 12, 22, 23].
Многочисленные исследования, проводимые по мониторингу возникновения новых штаммов B. рertussis, показали, что появляются штаммы B. pertussis с генетическими мутациями, изменяющими структуру протективных антигенов, способные уклоняться от иммунной защиты [14–23]. Мультилокусное антигенное сиквенстипирование (MAST) основано на изучении изменений в популяции B. pertussis. Схема генотипирования включает изучение последовательностей генов, кодирующих протективные антигены, в том числе входящих в состав современных бесклеточных вакцин: ptxA (кодирует S1-субъединицу коклюшного токсина), ptxВ (кодирует S2-субъединицу коклюшного токсина), ptxC (кодирует S3-субъединицу коклюшного токсина), prn (кодирует адгезивный белок пертактин), fim2 и fim3 (кодируют фимбриальные белки Fim2 и Fim3 соответственно). Промотор коклюшного токсина ptxP также включен для типирования изолятов, поскольку установлено, что аллель ptxP3 усиливает выработку токсина [25].
На территории РФ используют как цельноклеточную, так и бесклеточные коклюшные вакцины, входящие в состав комбинированных препаратов. Таким образом, целью данного исследования было изучение биологических и генотипических свойств штаммов B. pertussis, кандидатов в производственные вакцинные штаммы.
МАТЕРИАЛЫ И МЕТОДЫ
В рамках работы Референс-центра по мониторингу за коклюшем и дифтерией в Московский научноисследовательский институт эпидемиологии и микробиологии имени Г. Н. Габричевского Роспотребнадзора поступают бактериальные штаммы бордетелл и клинические образцы, полученные от больных коклюшем, согласно нормативным документам РФ, для верификации и генотипирования B. pertussis. Среди верифицированных штаммов нами было отобрано восемь штаммов B. pertussis в I фазе роста (гладкая форма) с хорошими ростовыми свойствами, которые были изучены согласно нормативным документам РФ по морфологическим, культуральным, ферментативным, серологическим и генотипическим свойствам с целью подбора кандидатных штаммов в состав коклюшных вакцин. Одним из критериев отбора было также выделение штаммов B. pertussis из различных регионов РФ.
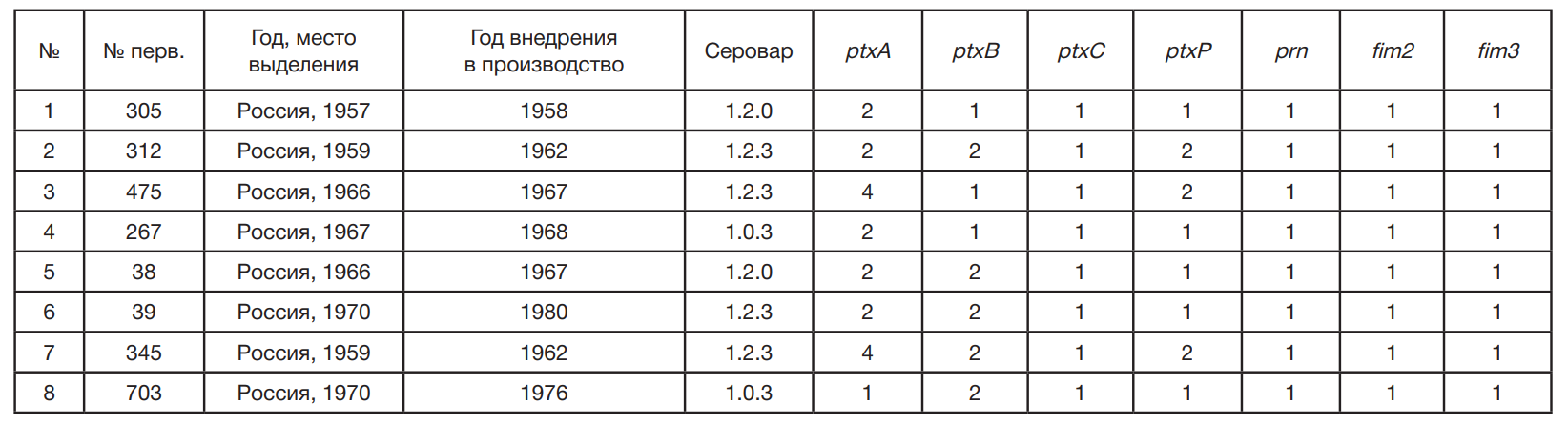
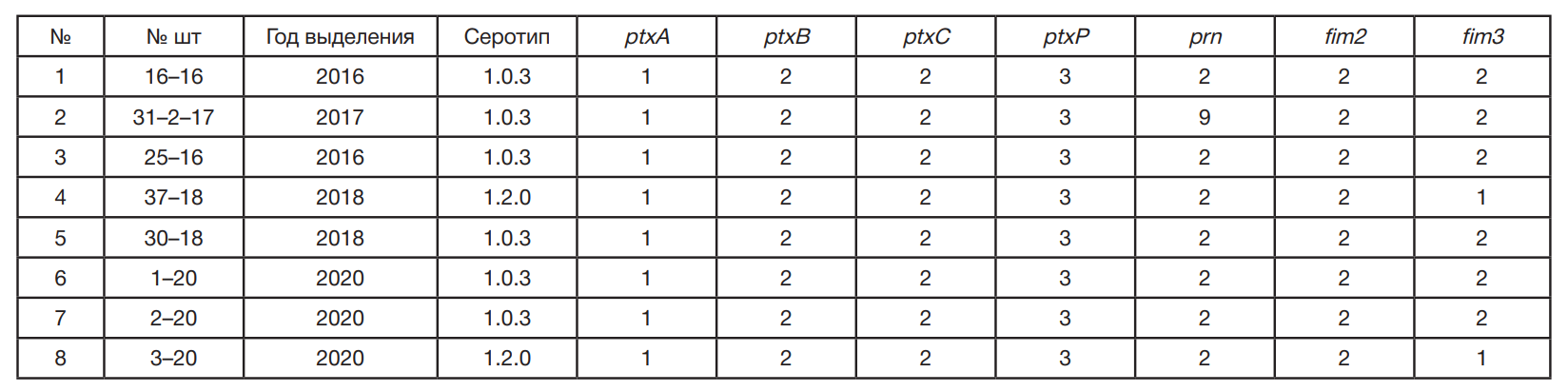
Штаммы B. pertussis были выделены от больных коклюшем разного возраста в период 2016–2020 гг. в г. Москве, Воронежской, Новосибирской, Ульяновской и Челябинской областях. Для сравнения использовали восемь вакцинных штаммов из коллекции ФГБУ «Научный центр экспертизы средств медицинского применения» Минздрава России (Москва), используемых в производстве коклюшного компонента АКДС-вакцины (табл. 1).
Культивирование штаммов B. pertussis проводили на плотной питательной среде Бордетелагар (ФБУН ГНЦ ПМБ, Оболенск, Россия) в течение 72 ч при + (36–37) °С. Идентификацию микроорганизмов проводили по культурально-морфологическим, тинкториальным и биохимическим свойствам. Культурально-морфологические свойства выросших колоний оценивали с помощью стереоскопического микроскопа SteREO Discovery V12 (Carl Zeiss; Германия) (объектив PlanApo S 1,0 × FWD 60 мм; окуляр PI 10 × 23 Br foc). Тинкториальные свойства изучали путем окраски по Граму («ЭКОлаб»; Россия). Окрашенные мазки просматривали с помощью светового микроскопа Axio Scope А1 (объектив EC Plan-NEOFLUAR 100 х 1,3; окуляр PI 10 × 23 Br foc (Carl Zeiss; Германия)).
Идентификацию штаммов B. pertussis по биохимическим свойствам проводили согласно нормативным документам [26].
Антигенную структуру (серотипы) штаммов оценивали в развернутой реакции агглютинации с использованием сывороток коклюшных к агглютиногенам 1,2,3, адсорбированных для реакции агглютинации (РА) сухих («НПО Микроген»; Россия).
Было проведено полногеномное секвенирование восьми производственных штаммов B. pertussis, используемых для производства коклюшного компонента АКДС-вакцины в настоящее время, и восьми штаммов B. pertussis, выбранных по микробиологическим и ростовым характеристикам как кандидатные. Генотипирование штаммов B. pertussis проводили по двум схемам: с использованием MAST путем анализа последовательности генов, кодирующих протективные антигены возбудителя — ptxA, ptxВ, ptxC, ptxP, prn, fim2 и fim3, согласно опубликованным рекомендациям [27] с использованием баз данных GenBank и BIGSdb, и полногеномного мультилокусного сиквенс-типирования (wgMLST).
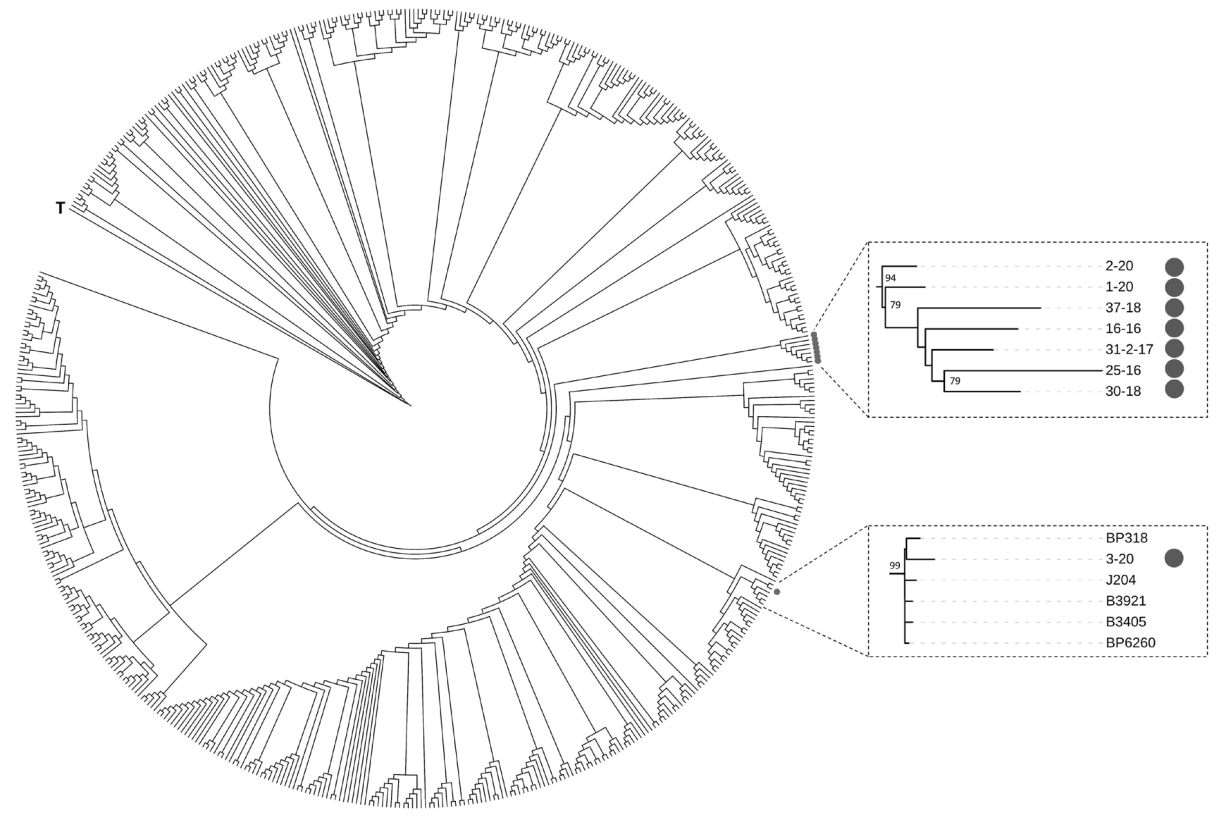
Выделение геномной ДНК из бактериальной культуры проводили с использованием набора «ExtractDNA Blood&Cells» («Евроген»; Россия). Полногеномное секвенирование осуществляли на платформе GenoLab (GeneMind Biosciences; Китай) с использованием наборов «SG GM» (Raissol; Россия) согласно рекомендациям производителя. Сборку осуществляли с помощью SPAdes-3.15.4, проверку качества сборки — с использованием QUAST 5.2.0 (https://github.com/ablab/quast; Россия). Выявление аллелей интересующих генов проводили на сервере BIGSdb. В качестве сравнения полученных нуклеотидных последовательностей при MAST использовали следующие номера референсных генов: ген ptxА (ptxА1 (AJ245366), ptxA2 (AJ245367), ptxA4 (AJ245368)); ген ptxВ (HM185483.1) (ptxВ1, ptxВ2); ген ptxC (AJ420987) (ptxC1 (M13223), ptxC2 (AJ420987)); промотор коклюшного токсина ptxP (ptxР1 (FN252323.1), ptxР2 (FN252322.1), ptxР3 (FN252324.1)); ген fim2 (fim2-1 (KT194049), fim2-2 (AJ420988)); ген fim3 (fim3-1 (X51543.1) и fim3-2 (AY464180.1)); ген prn (prn1 (AJ011091.1), prn2 (AJ011092.1), prn9 (AJ315611.1)). Для построения дерева на основе полногеномного мультилокусного сиквенс-типирования (wgMLST) поиск аллелей осуществляли программным обеспечением pyMLST 2.1.65 (https://github.com/bvalot/pyMLST/; Франция). В качестве штаммов сравнения использовали все полные геномы штаммов сиквенс-типа ST2, доступные в публичной базе данных NCBI Refseq, а также штамм Tohama I, относящийся к ST1, в качестве внешнего представителя. Для каждого штамма составляли аллельный профиль на основе последовательностей 2974 генов, после чего считали матрицу дистанций, отражающую количество несовпадающих аллелей между штаммами. На основе матрицы дистанций осуществляли построение дерева по алгоритму Neighbor-Joining в программном обеспечении rapidNJ 2.3.2 (https://github.com/somme89/rapidNJ; Дания).
Из описанного выше дерева были взяты две клады, одна из которых состояла целиком из штаммов-кандидатов, описанных в данной работе, а другая включала в себя штамм 3–20 и пять штаммов сравнения. Для этих штаммов (а также Tohama I в качестве внешнего представителя) были взяты последовательности всех 2974 исследуемых генов, конкатенированы и использованы для построения дерева по алгоритму Neighbor-Joining c использованием модели дистанций Кимуры. Данный подход был применен для оценки уровней поддержки bootstrap в описанных выше кладах предыдущего дерева.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
При культивировании на Бордетелагаре через 72 ч роста все восемь исследуемых штаммов B. pertussis вырастали в виде выпуклых круглых блестящих гладких колоний серовато-белого цвета размером до 1,5 мм, имели маслянистую консистенцию и легко снимались петлей. При просмотре в стереоскопическом микроскопе SteREO Discovery V12с объективом PlanApo S 1,0 × FWD 60 мм; окуляром PI 10 × 23 Br foc (Carl Zeiss; Германия) отмечали узкий луч света («хвостик»), отходящий от центра колонии. При микроскопии идентифицировали мелкие хаотично расположенные грамотрицательные палочки. Все исследуемые штаммы B. pertussis обладали каталазной и оксидазной активностью, не росли на кровяном и мясо-пептонном агаре, не продуцировали ферменты тирозиназу и уреазу, не росли на цитратном агаре Симмонса, не редуцировали нитраты в нитриты, не обладали подвижностью. При изучении антигенной структуры все исследуемые штаммы B. pertussis характеризовались присутствием агглютиногена 1 (видовой признак) и агглютинировались адсорбированными типоспецифическими сыворотками к агглютиногенам 1,2,3 не ниже 1 : 280. Производственные вакцинные штаммы относились к трем разным серотипам — 1.2.0, 1.0.3 и 1.2.3, штаммы–кандидаты к двум серотипам — 1.0.3 и 1.2.0 (табл. 1, табл. 2).
По последовательности гена ptxA, кодирующего S1-субъединицу коклюшного токсина, большинство производственных вакцинных штаммов B. pertussis соответствуют двум аллелям гена ptxА — ptxA4 и ptxA2, и только один производственный штамм B. pertussis, как и все современные кандидатные штаммы B. pertussis имеет ptxА1 аллель. Аллель ptxА1 отличается от других аллелей значимыми мутационными изменениями с заменой на аминокислотном уровне в Т-эпитопе S1-субъединицы коклюшного токсина (положения 204, 586, 668 и 969 нуклеотидной последовательности), имеющем непрерывную иммунодоминантную структуру, распознаваемую моноклональными (МкАТ) протективными m-антителами (mАТ). Аллель ptxА1 отличается от аллеля ptxA4 в положениях D68E, I228M и I232V и отличается от аллеля ptxA2 в положении I228M [12, 27].
По последовательности гена ptxВ, кодирующего S2-субъединицу коклюшного токсина, три производственных вакцинных штамма B. pertussis соответствуют аллелю ptxВ1 гена, большинство производственных штаммов (пять штаммов) и все современные кандидатные штаммы B. pertussis имеют аллель ptxВ2. Аллель ptxB1 отличается от аллеля ptxB2 изменением G18S в S2-субъединице коклюшного токсина.
При изучении фрагментов гена ptxC, кодирующего S3-субъединицу В-комплекса коклюшного токсина, выявлены штаммы двух вариантов последовательности нуклеотидов — ptxC1 и ptxC2. Все производственные вакцинные штаммы B. pertussis несли аллель ptxC1, в то время как все современные штаммы — кандидаты B. pertussis несли аллель ptxC2, который отличается от аллеля ptxC1 заменой нуклеотида в положении С681Т без изменений на аминокислотном уровне.
Хотя промотор коклюшного токсина ptxP не входит в компоненты бесклеточной вакцины, его обычно используют при типировании штаммов в качестве маркера возникшей генетической линии, которая распространилась по всему миру [14–23, 27]. Изучение последовательности промоторной области ptxP коклюшного токсина показало, что у изученных штаммов присутствуют три аллеля ptxР — ptxР1, ptxР2 и ptxР3. Пять производственных вакцинных штаммов B. pertussis несли аллель ptxP1 и три производственных штамма — аллель ptxР2. Все кандидатные штаммы B. pertussis имели аллель ptxP3.
Аллель ptxР3 отличается от ptxР1 и ptxР2 мутацией в положении –65 нуклеотидной последовательности, что обеспечивает повышенную прочность связывания с димером BvgA, приводящую к увеличению продукции коклюшного токсина [25].
При секвенировании гена fim3, кодирующего фимбриальный белок Fim3, идентифицировано два варианта нуклеотидной последовательности — fim3-1 и fim3-2. У всех производственных вакцинных штаммов идентифицирован одинаковый аллель fim3-1, среди кандидатных штаммов B. pertussis у большинства штаммов (шесть штаммов) выявлен аллель fim3-2 и у двух штаммов — fim3-1. Последовательность нуклеотидов аллеля fim3-2 отличается от последовательности нуклеотидов аллеля fim3-1 значимой мутацией, которая сопровождается изменениями на аминокислотном уровне A87E в молекуле белка Fim3.
При секвенировании гена fim2, кодирующего фимбриальный белок Fim2, идентифицировано два варианта нуклеотидной последовательности: fim2-1 и fim2-2. У всех производственных вакцинных штаммов выявлен одинаковый аллель fim2-1, у всех кандидатных штаммов B. pertussis — fim2-2. Выявленные аллельные варианты fim2-1 и fim2-2 различались изменениями R174K в молекуле белка Fim2.
Ген prn содержит около 2800 оснований, которые кодируют крупный адгезивный белок предшественник — 5'-конец гена кодирует часть предшественника пертактина, которая экпортируется из клетки, в то время как 3'-конец кодирует интегральный белок внешней мембраны, необходимый для этого экспорта. При секвенировании гена пертактина (prn) у изученных штаммов B. pertussis выявлено три варианта аллелей гена prn — prn1, prn2 и prn9. Среди производственных вакцинных штаммов идентифицирован аллель prn1. У кандидатных штаммов B. pertussis идентифицированы prn2 (у семи штаммов) и prn9 (у одного штамма). Последовательности нуклеотидов аллелей prn2 и prn9 отличаются от последовательности у аллеля prn1 наличием значимых мутационных изменений в шести положениях (828, 831, 832, 833, 834 и 836) нуклеотидной последовательности гена prn с заменами на аминокислотном уровне — V279G и A278F. Последовательность нуклеотидов аллеля prn2 имеет вставку дополнительного фрагмента в 15 п.н. — GGCGGCCTCGGTCCC в 1-й области гена в положениях 841–855, а prn9 аллель гена — дополнительный фрагмент в 30 п.н. — GGCGGCCTCGGTCCCGGCGGCCTCGGTCCC в 1-й области гена в положениях 841–871. Все изменения в prn2 и prn9 аллелях произошли в 1-й и 2-й областях Prn белка, которые иммуногенны и участвуют в формировании В-клеточного иммунного ответа.
В результате проведенного исследования выявлено, что все производственные вакцинные штаммы принадлежали к шести генотипам — ptxА2/ptxВ1/ptxС1/ptxР1/fim2-1/fim31/prn1, ptxА2/ptxВ2/ptxС1/ptxР2/fim2-1/fim3-1/prn1, ptxА4/ptxВ1/ptxС1/ptxР2/fim2-1/fim3-1/prn1, ptxА2/ptxВ2/ptxС1/ ptxР1/fim2-1/fim3-1/prn1, ptxА4/ptxВ2/ptxС1/ptxР2/fim2-1/fim3-1/prn1 и ptxА1/ptxВ2/ptxС1/ptxР1/fim2-1/fim3-1/prn1 (табл. 1). Кандидатные штаммы являются представителями четырех генотипов — ptxА1/ptxВ2/ptxС2/ptxР3/fim2-2/fim3-2/prn2, ptxА1/ptxВ2/ptxС2/ptxР3/fim2-2/fim3-2/prn9, ptxА1/ptxВ2/ptxС2/ptxР3/fim2-1/fim3-1/prn1 и ptxА1/ptxВ2/ptxС2/ ptxР3/fim2-2/fim3-1/prn2 (табл. 2).
На основании последовательностей полных геномов кандидатных штаммов было реконструировано филогенетическое дерево, отражающее эволюционное положение исследованных штаммов-кандидатов среди всех представителей сиквенс-типа ST2 (рисунок). Установлено, что все из них, кроме одного, формируют единый близкородственный кластер, по-видимому, характерный для территории РФ.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Все полученные в ходе исследования геномные последовательности штаммов B. pertussis включены в Национальный каталог на базе ФБУН ГНЦПМБ Роспотребнадзора в рамках федерального проекта «Санитарный щит страны — безопасность для здоровья (предупреждение, выявление, реагирование)».
При многолетнем мониторинге генотипических свойств B. pertussis установлено, что за период более чем 60-летней плановой иммунизации детского населения в популяции штаммов возбудителя коклюша распространяются штаммы с новыми генотипами (аллельными профилями) [14–16], что полностью соответствует общемировой тенденции изменения структуры генов протективных антигенов B. pertussis [18–23]. Исследования, проводимые в разных странах мира, также показали генотипические различия между штаммами, используемыми для производства вакцинных препаратов, и циркулирующей популяцией возбудителя коклюша [11, 18–23, 28, 29].
В нашем исследовании охарактеризованы и предложены в качестве кандидатных восемь штаммов B. pertussis, относящихся к четырем разным генотипам, отличающихся от генотипов производственных вакцинных штаммов, используемых для получения АКДС-вакцины в настоящее время. Для включения штаммов-кандидатов B. pertussis в состав банка производственных вакцинных штаммов в дальнейшем будут изучены их иммунобиологические свойства с определением гемагглютинирующей, гемолитической и лейкоцитозстимулирующей активностей, вирулентности и токсичности на животных, в также проведено депонирование в Государственной коллекции патогенных микроорганизмов («ГКПМ-Оболенск»).
В связи с тем что изменения циркулирующих штаммов B. pertussis идут непрерывно, мониторинг генетических свойств штаммов должен проходить постоянно, что дает возможность оценивать влияние бактериальных изменений на эффективность вакцины. Следует продолжить проведение исследований по изучению генотипа циркулирующих штаммов B. pertussis и особенностей формирования в современных условиях постинфекционного и поствакцинального иммунного ответа — для обоснованной оценки адекватности используемых цельноклеточных и бесклеточных коклюшных вакцин как средства профилактики коклюшной инфекции. Для оценки протективности производимых вакцин необходимы также модельные опыты на животных с использованием штаммов B.рertussis современных генотипов.
ВЫВОДЫ
В результате проведенного исследования подобраны штаммы B. рertussis в качестве кандидатов для производства коклюшных вакцин с оценкой ростовых, культурально-морфологических и генотипических свойств.
- Петрова М. С., Попова О. П., Борисова О. Ю., Абрамова Е. Н., Вартанян Р. В., Келли Е. И. Коклюш у детей раннего возраста. Эпидемиология и инфекционные болезни. 2012; 6: 19–24. DOI: 10.17816/EID40704.
- Бабаченко И. В., Нестерова Ю. В., Чернышова Ю. Ю., Карасев В. В., Починяева Л. М., Калисникова Е. Л. Клиникоэпидемиологические аспекты коклюша у детей в условиях массовой вакцинопрофилактики. Журнал инфектологии. 2019; 11 (2): 88–96. DOI: 10.22625/2072-6732-2019-11-2-88-96.
- Краснов В. В., Ильяненков К. Ф., Павлович Л. Р., Кузмичева М. В. Коклюш у детей первого года жизни. Детские инфекции. 2018; 1 (1): 12–17. DOI: 10.22627/2072-8107-2018-17-1-12-17.
- Об организации проведения внешнего контроля качества исследований по диагностике дифтерии и коклюша в 2024 году в Дальневосточном и Уральском федеральных округах. Информационное письмо № 02/1860-2024-27 от 06.02.2024. Федеральная служба по надзору в сфере защиты прав потребителей и благополучия человека (Роспотребнадзор).
- Басов А. А., Высочанская С. О., Цвиркун О. В., Белова Т. Р., Адугюзелов С. Э., и др. Критерии оценки эпидемиологической ситуации по коклюшу в Российской Федерации. Эпидемиология и вакцинопрофилактика. 2024; 23 (2): 4–13. DOI: 10.31631/2073-3046-2024-23-1-4-13.
- О состоянии санитарно-эпидемиологического благополучия населения в Российской Федерации в 2021 году: Государственный доклад. М.: Федеральная служба по надзору в сфере защиты прав потребителей и благополучия человека, 2022; 340 с.
- Нестерова Ю. В., Медкова А. Ю., Бабаченко И. В., Семин Е. Г., Калисникова Е. Л., Синяшина Л. Н. Клинико-диагностическое значение генетических маркеров Bordetella pertussis у контактных лиц в семейных очагах. Журнал инфектологии. 2019; 11 (1): 17–24. DOI: 10.22625/2072-6732-2019-11-1-17-24.
- Philipson K, Goodyear-Smith F, Grant CC, Chong A, Turner N, Stewart J. When is acute persistent cough in school-age children and adults whooping cough? British Journal of General Practice. 2013; 63 (613): 573–9. DOI: 10.3399/bjgp13X670705.
- Пименова А. С., Борисова О. Ю., Цвиркун О. В., Басов А. А., Алешкин В. А., Афанасьев С. С., и др. Эффективность применения молекулярно-генетической диагностики при обследовании очагов коклюшной инфекции. Инфекция и иммунитет. 2017; 7 (2): 162–70. DOI: 10.15789/2220-7619-2017-2-162-170.
- Скирда Т. А., Борисова О. Ю., Петрова М. С., Борисова А. Б. Серологическая диагностика коклюша у лиц старшего возраста. Клиническая лабораторная диагностика. 2020; 65 (8): 492–5. DOI: 10.18821/0869-2084-2020-65-8-492-495.
- Esposito S, Stefanelli P, Fry NK, Fedele G, He Q, Paterson P, et al. Pertussis prevention: reasons for resurgence, and differences in the current acellular pertussis vaccines. Frontiers in Immunology. 2019; 10: 1344. DOI: 10.3389/fimmu.2019.01344.
- Mooi FR, Van Der Maas NA, De Melker HE. Pertussis resurgence: waning immunity and pathogen adaptation — two sides of the same coin. Epidemiol Infect. 2014; 142 (4): 685–94. DOI: 10.1017/S0950268813000071.
- Скирда Т. А., Борисова О. Ю., Борисова А. Б., Комбарова С. Ю., Пименова А. С., Гадуа Н. Т., и др. Определение противококлюшных антител у школьников с длительным кашлем. Журнал инфектологии. 2023; 15 (1): 93–100. DOI: 10.22625/2072-6732-2023-15-1-93-100.
- Борисова О. Ю., Гадуа Н. Т., Пименова А. С., Петрова М. С., Попова О. П., Алешкин В. А., и др. Структура популяции штаммов возбудителя коклюша на территории России. Эпидемиология и вакцинопрофилактика. 2016; 4: 22–28. DOI: 10.31631/2073-3046-2016-15-4-22-28.
- Андриевская И. Ю., Борисова О. Ю., Пименова А. С., Борисова А. Б., Гадуа Н. Т., Чагина И. А. Генотипирование возбудителя коклюша из клинических образцов. Материалы XVI Ежегодного Всероссийского конгресса по инфекционным болезням имени академика В.И. Покровского к 300-летию Российской академии наук, Москва, 25–27 марта 2024 г. М.: Медицинское маркетинговое агентство, 2024; 250 с: 19–20. ISBN 978-5-6048391-2-6.
- Андриевская И. Ю., Пименова А. С., Гадуа Н. Т., Чагина И. А., Борисова О. Ю. Аллельное типирование ptxP Bordetella pertussis в клинических образцах. Материалы конгресса с международным участием «Молекулярная диагностика и биобезопасность – 2024», Москва, 16 – 17 апреля 2024 г. М.: ФБУН ЦНИИ Эпидемиологии Роспотребнадзора, 2024; 266 с.: 36. DOI: 10.36233/978-5-6048873-9-4.
- Kentaro K, ShuMan Y, Chuen-Sheue C, Phung TBT, Do TTN, et al. Genotyping and macrolide-resistant mutation of Bordetella pertussis in East and South-East Asia. J Glob Antimicrob Resist. 2022; 31: 263–9. DOI: 10.1016/j.jgar.2022.10.007.
- Barkoff AM, Mertsola J, Pierard D, Dalby T, Hoegh SV, Guillot S, et al. Surveillance of Circulating Bordetella pertussis Strains in Europe during 1998 to 2015. J Clin Microbiol. 2018; 56 (5): e01998-17. DOI: 10.1128/JCM.01998-17.
- Moosa F, du Plessis M, Weigand MR, Peng Y, Mogale D, de Gouveia L, et al. Genomic characterization of Bordetella pertussis in South Africa, 2015–2019. Microb Genom. 2023; 9 (12): 001162. DOI: 10.1099/mgen.0.001162.
- Fu P, Zhou J, Yang C, Nijiati Y, Zhou L, Yan G, et al. Molecular Evolution and Increasing Macrolide Resistance of Bordetella pertussis, Shanghai, China, 2016–2022. Emerg Infect Dis. 2023; 30 (1): 29–38. DOI: 10.3201/eid3001.221588.
- Weigand MR, Williams MM, Peng Y, Kania D, Pawloski LC, Tondella ML. Genomic Survey of Bordetella pertussis Diversity, United States, 2000–2013. Emerg Infect Dis. 2019; 25 (4): 780–3. DOI: 10.3201/eid2504.180812.
- Zomer A, Otsuka N, Hiramatsu Y, Kamachi K, Nishimura N, Ozaki T, et al. Bordetella pertussis population dynamics and phylogeny in Japan after adoption of acellular pertussis vaccines. Microb Genom. 2018; 4 (5): 000180. DOI: 10.1099/mgen.0.000180
- Mir-Cros A, Moreno-Mingorance A, Martín-Gómez MT, Codina G, Cornejo-Sánchez T, Rajadell M, et al. Population dynamics and antigenic drift of Bordetella pertussis following whole cell vaccine replacement, Barcelona, Spain, 1986– 2015. Emerg Microbes Infect. 2019; 8 (1): 1711–20. DOI: 10.1080/22221751.2019.1694395.
- Althouse BM, Scarpino SV. Asymptomatic transmission and the resurgence of Bordetella pertussis. BMC Med. 2015; 13: 146. DOI: 10.1186/s12916-015-0382-8.
- Mooi FR, van Loo IH, van Gent M, He Q, Bart MJ, Heuvelman KJ, et al. Bordetella pertussis strains with increased toxin production associated with pertussis resurgence. Emerg Infect Dis. 2009; 15 (8): 1206–13. DOI: 10.3201/eid1508.081511.
- Методические указания «Лабораторная диагностика коклюша и заболеваний, обусловленных другими бордетеллами». МУК 4.2.3701-21. М.: Федеральная служба по надзору в сфере защиты прав потребителей и благополучия человека. 2021.
- Mooi FR, Hallander H, Wirsing von König CH, Hoet B, Guiso N. Epidemiological Typing of Bordetella pertussis Isolates: Recommendations for a Standard Methodology. Eur J Clin Microbiol Infect Dis. 2000; 19: 174–81.
- Gordana D, Teemu K, Annika E, Pljesa T, Vignjevic-Krastavcevic M, He Q. Bordetella pertussis vaccine strains and circulating isolates in Serbia Vaccine. 2010; 28 (5): 1188–92. DOI: 10.1016/j.vaccine.2009.11.036.
- Alai S, Ghattargi VC, Gautam M, Patel K, Pawar SP, Dhotre DP, et al. Comparative genomics of whole-cell pertussis vaccine strains from India. BMC Genomics. 2020; 21 (1): 345. DOI: 10.1186/s12864-020-6724-8.